Those keeping up with the pterosaurian literature will be
aware that the latest issue of Acta Geologica Sinica has a set of papers
resulting (at least in part) from the 2010 Beijing Flugsaurier meeting. (And
while we’re on the subject, the 2013 meeting in Rio has extended abstract
submissions till the 31st of January, so there’s still time to get
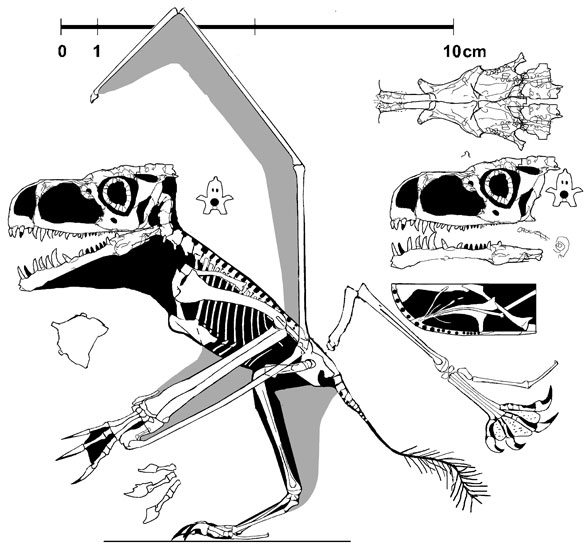
them in). I’ve got a couple in there and while people might be more interested
in the horribly flattened anuroganthid, I’m more keen to talk about the short
review paper I produced.
The title, ‘Pterosaur Research: Recent Advances and a Future
Revolution’, might sound familiar and indeed some of the content may too.
That’s because it ultimately sprung out of a post that I had over on the
Musings and also put up on Pterosaur.net. This is a first for me at least, a
paper that resulted pretty much directly from a blogpost.
At the time I’d been writing about rates of discovery of
dinosaurs and pterosaurs and what that might mean for future discoveries. It
occurred to me that actually the pterosaurs seemed to be going through
something of a renaissance in the way that dinosaurs had in the 1970s. We were
finding more and more of them, more papers were being published by more
researchers, and more of the big questions were either being answered, or at
least were being tackled in a rather more systematic way than they had before.
It occurred to me that this was worth summarising and producing something more
formal. The fact that the next Flugsaurier volume was due meant there was a
most suitable venue available and discussions with various colleagues helped me
develop the idea and push for it’s inclusion.
If you look back at the developments of the last 10 years or
so in pterosaurs, it’s quite a remarkable and rapid progression. That’s not to
overlook the huge amount of groundwork that had gone before and the efforts of
previous generations, but even quite a few fundamentals that had occurred for
dinosaurs decades ago are now being sorted out for pterosaurs. We now have
inclusive phylogenies for pterosaurs, we’ve got a good idea of their soft
tissue structures and especially the wing, some of the taxonomic and systematics
issues of the past are being resolved, we’ve got a major transition in the form
of Darwinopterus, cool new taxa like the boreopterids and chaoyangopterids turning up, detailed
analyses of flight, take-off, mass estimates, muscle patterns, and skull
shapes, we finally, finally, have eggs and we’re even getting serious on
behaviour and ecology for analyses of head crests, growth and the like as well
as looking at major evolutionary trends like diversification and distribution.
We’re even getting attention from the public and serious attention with whole
exhibits on pterosaurs, new books, and documentaries, and of course we now have
the Flugsaurier meetings themselves, established and (hopefully) regular events
that will help keep things ticking along.
So this paper attempts to summarise all of this and in
effect provide a statement of the ‘state of the art’ – what do we know and how
have we got there, but it is also supposed to be a bit of a celebration of the
last decade of research and the gains made by the pterosaur research community.
Those in the know will probably realise that the background to this has not
been without a significant amount of strife, and while this is not mentioned in
the paper, I think it only emphasises how much has been learned despite this
limitation. I hope it also provides a sort of counter-point, but also a
continuation of Peter Wellnhofer’s piece that kicked off the 2008 Flugsauriervolume. Peter wrote a review of the history of pterosaur research, but pretty
much only took it up to the modern era, and with the galloping developments of
the last few years, this should bring things more or less up to date.
The paper is available here (and indeed all of the latest
papers are from AGS). Just click on the left hand set of the three series of
Chinese characters at the bottom of the page and then add a .pdf suffix to the
filename once it’s been saved.













{kind=link}