
It's a common misconception that staking the heart of a Stokerian vampire will do it in for good. In actuality, the characters of Bram Stoker's Dracula (above) only considered the undead truly out of action once they were staked through the heart (which only imobilised them, see, not killing them), decapitated, had their mouths stuffed with garlic flowers and the access to their tombs lined with holy masonry. This may seem like overkill, but, for the vampire mythos Stoker created, it is the only way to keep the blaggards down. In the last week, it's emerged that that palaeontological vampires need a similar heavy duty approach to ensure that they too don't continue to rise from the grave.
 Enter, stage left, the hypothesis that the anuroganthid Jeholopterus was a Mesozoic vampire bat equivalent (image, above, from my upcoming book, shows the anuroganthid Anurognathus with a more accurate Insect Hawking Cookie Monster of Doom appearance, not a vampire). Proposed by David Peters in an abstract for the SVP annual conference of 2003, cited evidence for this idea stems from large caniform teeth inferred on the Jeholopterus holotype using DP's infamous digital photo interpretation (for anyone unfamilar with DP's work, you can see the most recent incarnation of it here). It is well known that palaeontologists have almost never seen eye to eye with Peters' interpretations of fossils or methods of analysing them, and a small body of literature exists that directly refutes his work (e.g. Bennett 2005; Hone et al. 2009) . Many other papers also disagree with his methods or conclusions. DP acknowledges his 'heretical' views and, indeed, has even named his blog after them: The Pterosaur Heresies. Our very own Pterosaur.Net even gets a good kicking at various points at TPH, but that's OK: we have our opinions (which we consider to be well supported and credible), and Dave has his (which we consider to be very poorly supported). I think we have to live with the fact that we're not going to agree with everyone in science, and, frustrating though this can be at times, we're much better off making sure our own work is as watertight as possible than constantly bickering with others.
Enter, stage left, the hypothesis that the anuroganthid Jeholopterus was a Mesozoic vampire bat equivalent (image, above, from my upcoming book, shows the anuroganthid Anurognathus with a more accurate Insect Hawking Cookie Monster of Doom appearance, not a vampire). Proposed by David Peters in an abstract for the SVP annual conference of 2003, cited evidence for this idea stems from large caniform teeth inferred on the Jeholopterus holotype using DP's infamous digital photo interpretation (for anyone unfamilar with DP's work, you can see the most recent incarnation of it here). It is well known that palaeontologists have almost never seen eye to eye with Peters' interpretations of fossils or methods of analysing them, and a small body of literature exists that directly refutes his work (e.g. Bennett 2005; Hone et al. 2009) . Many other papers also disagree with his methods or conclusions. DP acknowledges his 'heretical' views and, indeed, has even named his blog after them: The Pterosaur Heresies. Our very own Pterosaur.Net even gets a good kicking at various points at TPH, but that's OK: we have our opinions (which we consider to be well supported and credible), and Dave has his (which we consider to be very poorly supported). I think we have to live with the fact that we're not going to agree with everyone in science, and, frustrating though this can be at times, we're much better off making sure our own work is as watertight as possible than constantly bickering with others.The vampire Jeholopterus made a brief splash back in 2003, but was widely condemned by the entire pterosaur community. To many, this simply proved - again - that SVP perhaps needed to pay closer attention to the work they were allowing into their conference, but that was that. No peer reviewed paper on the vampire hypothesis followed, and no independent confirmation that Jeholopterus or other anurognathids were sanguivorous has been proposed. Instead, the long-held view that anurognathids were ace aerial insect hawkers has prevailed (e.g. Bennett 2007; Habib 2011 [a follow up publication to which is in the works. I'm lucky enough to have been invited in on the authorship and can promise that some of the stuff in it should blow your little socks off]). The vampire pterosaur idea, it seemed, was dead, the only remnants being the abstract, a few media stories, and the Jeholopterus page at DP's website. This week, however, the vampire Jeholopterus meme has risen from the grave, being portrayed in a half-credible light in this article and picked up elsewhere online. Several people, including myself, were a bit miffed at this, and, in full on SIWOTI mode, left comments on these articles. The original article seems to be picking comments that agree with the tone of the article as their 'featured' comments, hiding perhaps more informed opinions in other pages of the article. Hence, seeing as most people won't easily find these remarks, I thought best to regurgitate mine here. In short, I want to provide a one-stop shop for clarity on the vampire pterosaur hypothesis:
- The idea was not peer reviewed, and it's publication in a collection of conference abstracts is not of comparable standing to other hypotheses of anurognathid palaeoecology
- There was never any 'debate' amongst pterosaur workers on this idea: it was never considered credible by qualified researchers in the first instance, and rejected outright from the start.
- There is no evidence that Jeholopterus, or any other pterosaur, was a vampire
- There is no 'David Peters vs. Goliath' story here. DP's work is considered with the same scrutiny, not more or less, than any other piece of science. His ideas are rejected by other palaeontologists (amateur and professional alike, the only difference between many of whom is that some are paid to study fossils) because they have not stood up to this scrutiny.
--
This story has been told rather incorrectly. DP's 'publication' in 'the peer-reviewed Journal of Vertebrate Paleontology' was NOT peer reviewed: it was a short abstract for the SVP 2003 annual conference. I am confident as a 'professional' pterosaur worker myself that this paper would not have made it into any scientific journal, and it was rightly condemned by the pterosaur community as soon as it was made public. Along with the Bennett article mentioned here, a body of literature exists demonstrating that most, if not all, of David Peter's methods of reconstruction and image interpretation are flawed. The extraneous features he reconstructs for fossil animals (which have included, at one time or another, fantastic frills, sails, additional bones and teeth, long tails on short-tailed taxa, hatchlings clinging to their parent's body and others) have never been found on fossil specimens despite CT scanning, UV investigation and other analytical methods. The vast majority of DP's ideas are not corroborated by any studies except his own. In polite terms, DP's ideas are considered 'fringe' at best by palaeontologists, and very much the view of one individual. (animated vampire Jeholopterus feeding strategy, below)
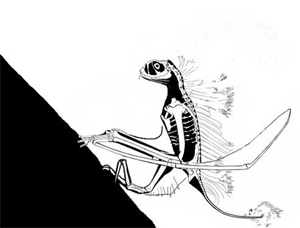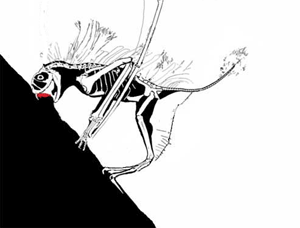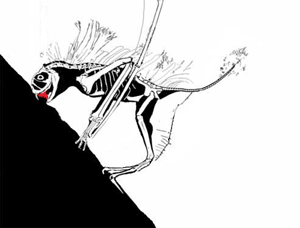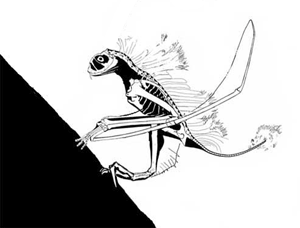
I find it worrying that you wrote your article without uncovering or featuring these details. Likewise, the fact that you give the vampire Jeholopterus idea some credence with statements like 'what spurred the great debate' and 'without a living Jeholopterus to observe, we really cannot be sure of its unique attributes': there was never any debate, and the latter suggests a critical misunderstanding of scientific practise. Palaeontologists work, like all other sciences, by testing hypotheses: we are confident that Jeholopterus was not a vampire bat-like animal because it fails tests we can put against this idea. Does it bear large teeth for piercing flesh? No. None have ever been found on any actual specimen: the fact they have been found on someone's computer screen means nothing if they cannot be seen by some means on the actual fossil. DP probably picked up compression artefacts in the jpeg or cracks, shadows and prep marks in the matrix on the slab. Did Jeholopterus have a strong bite? Probably not, as the bones of the jaw are mechanically weak and slender, and ill-suited to anchoring strong muscles. Are there any alternative means it could use to pursue a vampire lifestyle? None that we can ascertain. Is there a more plausible hypothesis for the lifestyle of Jeholopterus? Yes: aerial insectivory, a lifestyle that decades of _actual_ peer-reviewed studies into anurognathid (the group that Jeholopterus belongs to) anatomy and biomechanics support without exception.
Finally, the portrayal of DP as a maverick, lone amateur 'informing' the body of professional palaeontologists is unfair. A great number of so-called 'amateur' palaeontologists produce work of the highest credibility without a whiff of controversy. Like DP, they work alone and draw their own conclusions, but find that their ideas are similar to those lucky enough to be paid to research palaeontological subjects. There is no conspiracy about preservation of dogmatic ideas or rejection of outsiders: the internet teems with blogs and forums where paid palaeontologists and 'amateuers' meet to discuss ideas at the highest level. I'm afraid to say I find this article very ill-informed and misleading, and hope this comment adds some balance to this page.
References
- Bennett, S. C. 2005. Pterosaur science or pterosaur fantasy? Prehistoric Times, 70, 21-23.
- Bennett, S. C. 2007. A second specimen of the pterosaur Anurognathus ammoni. Paläontologische Zeitschrift, 81, 376-398.
- Habib, M. B. 2011. Functional morphology of anurognathid pterosaurs. Geological Society of America Abstracts with Programs, 43, 118.
- Hone, D. W. E., Sullivan, C. and Bennett S. C. 2009. Interpreting the autopodia of tetrapods: interphalangeal lines hinge on too many assumptions. Historical Biology, 21, 67-77.
- Peters, D. 2003. The Chinese vampire and other overlooked pterosaur ptreasures. Journal of Vertebrate Paleontology, 23(3), 87A.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}